25.1 INTRODUCTION
Impedance plethysmography is a method of determining changing tissue volumes in the body, based on the measurement of electric impedance at the body surface. This chapter presents the bioelectric basis of impedance plethysmography with emphasis on impedance cardiography - that is, determination of cardiac stroke volume. The first publications concerning this method date back to the 1930s and 1940s (Atzler and Lehmann, 1931; Rosa, 1940; Holzer, Polzer, and Marko, 1946; Nyboer et al., 1940; Nyboer, Bango, and Nims, 1943; Nyboer, 1950). The method reached clinical value about 20 years ago based on the research work by Kinnen, Kubicek, et al. (Kinnen et al., 1964,a,b,c; Kubicek et al., 1966; Kubicek, Patterson, and Witsoe, 1970). A related method, integral rheography, for measuring the cardiac output was developed by Ti
where all variables are evaluated at t0. Equation 25.2 describes how the macroscopic resistivity Z (impedance per unit volume) is derived from the spatial distribution of conductivity s weighted by the dot product of the lead fields of the current and voltage electrodes. Note the similarity between Equation 25.2 and the fundamental equation of the lead field theory, Equation 11.30 (or 11.52), which describes the electric signal in the lead produced by a volume source formed by a distribution of the impressed current
25.2.2 Tissue Impedance
The physical quantity measured in impedance plethysmography (and imaged in impedance tomography) is tissue impedance. (The impedance of various tissues was discussed in Section 7.4.) From Table 7.3 it can be seen that the resistivity of body organs varies about 100-fold from about 1.6 Wm in blood to about 170 Wm in bone. Within the soft tissues the variability is about 10-fold, with about 20 Wm in the lung and in fat.
The right-hand side of Equation 25.4 is a constant where one recognizes the equation to be that of a circle whose center is at Im = 0, Re = (R 0 - R
In the corresponding Cole-Cole plot, shown in Figure 25.1C, the depression angle is f = (1 - a)p/2. Figure 25.2 shows the depression of the semicircle in the Cole-Cole plots for the transverse and longitudinal impedances of skeletal muscle as measured by Epstein and Foster (1983).
 enko and coworkers (1973). This method has, however, hardly been used outside the Soviet Union.
enko and coworkers (1973). This method has, however, hardly been used outside the Soviet Union.
 Determination of the cardiac stroke volume is an area in which accurate, easily applied, noninvasive methods are needed. Impedance cardiography is easy to apply, noninvasive, and also cheap; however, it has serious methodological limitations, which are discussed below. We also provide a brief overview of other applications of impedance plethysmography.
The magnetic method corresponding to electric impedance plethysmography is called magnetic susceptibility plethysmography. This method may be used for monitoring blood volume changes in the thorax. Most living tissues are diamagnetic since water is their major constituent. If a strong magnetic field is applied to the region of the thorax, the movements of the heart, blood, and chest wall during the heart cycle cause variations in magnetic flux. Thus it is possible to monitor these variations with a SQUID magnetometer during the heart cycle (Wikswo, 1975; Maniewski et al., 1988). Currently, magnetic susceptibility plethysmography does not have clinical applications and, therefore, this method is not discussed in detail in this book.
Determination of the cardiac stroke volume is an area in which accurate, easily applied, noninvasive methods are needed. Impedance cardiography is easy to apply, noninvasive, and also cheap; however, it has serious methodological limitations, which are discussed below. We also provide a brief overview of other applications of impedance plethysmography.
The magnetic method corresponding to electric impedance plethysmography is called magnetic susceptibility plethysmography. This method may be used for monitoring blood volume changes in the thorax. Most living tissues are diamagnetic since water is their major constituent. If a strong magnetic field is applied to the region of the thorax, the movements of the heart, blood, and chest wall during the heart cycle cause variations in magnetic flux. Thus it is possible to monitor these variations with a SQUID magnetometer during the heart cycle (Wikswo, 1975; Maniewski et al., 1988). Currently, magnetic susceptibility plethysmography does not have clinical applications and, therefore, this method is not discussed in detail in this book.


(25.1)
where DZ = impedance change [W/mł] t0, t1 = time instants Ds = conductivity change between the two time instants [S/m = 1/W·m]  LE
LE = lead field of the voltage measurement electrodes for unit reciprocal current [1/m2] LI = lead field of the current feeding electrodes for unit current [1/m2] v = volume [m3] In Equation 25.1, the region v consists of an inhomogeneous volume conductor whose conductivity (as a function of position) at time t0 is s(t0). At t1, this has changed to s(t1), and it is this change (t1) - (t0) = Ds which is responsible for the measured impedance change DZ. Thus Equation 25.1 describes how the changes in volume conductor conductivity are converted into the impedance change evaluated from a measured voltage (at the voltage electrode pair) divided by applied current (at the current electrode pair). Note that the 4-electrode impedance method underlies Equation 25.1.
A special case of Equation 25.1 is one where we consider s(t1) = es(t0), where e is very small:

(25.2) i. In these equations the corresponding variables are the measured signals: VLE and Z (= measured voltage per applied current), the distributions of sensitivity: LE in both of them, as well as the source distributions: i and LI.
If the introduction of the current is done with the same electrodes as the voltage measurement is made, the sensitivity distribution, that is the lead field LE is the same as the distribution of the applied current LI. This technique is, however, seldom used because of the artifact due to the electrode impedance. If the current-feeding electrodes are different from those of the voltage measurement electrodes, the sensitivity distribution is the dot product of the lead fields of the voltage electrodes LE and the current electrodes LI. Thus, any previous discussion in this book on the electric and magnetic lead fields in general (Chapters 11 and 12), in the head (Chapters 13 and 14) or in the thorax (Chapters 15 ... 18 and 20) may readily be applied to impedance plethysmography. Just as in the study of electrocardiography, one can design electrode systems for impedance measurement to give special emphasis to particular regions (the aorta, the ventricles, etc.). One can even have situations where the dot product is negative in a particular region so that if the conductivity increases in that region, the impedance Z will also increase. Some examples can be found in Plonsey and Collin (1977) and Penney (1986).
While Equation 25.1 is a suitable theoretical basis for impedance plethysmography, we are still left with considerable uncertainty how varies throughout the heart and torso or in what way the circulation modifies the thorax structure and conductivity as a function of time throughout the cardiac cycle. Further research is required to develop a physiologically adequate circulation model. Note, however, that Equation 25.1 may be more readily applied over a longer time frame (t1 - t0) to, say, the growth of a localized tumor in the thorax (other regions remaining the same).
In measuring bioelectric sources the reactive component of tissue impedance is not important because the frequency range is under 1 kHz. Actually, in Section 7.2.4 it was shown that it can be omitted with the assumption of quasistationarity. In impedance plethysmography (and tomography) the frequency dependence of tissue impedance is a factor which can be utilized for increasing the selectivity of the system. Because the impedance of different tissues has different reactive components, the impedance may be measured with applied currents at different frequencies (Lozano, Rosell, and Pallás-Areny, 1990). The frequencies may be selected so that the separation of certain tissues is maximized. With appropriate filtering the measurement may be done simultaneously with different frequencies in order to save measurement time.
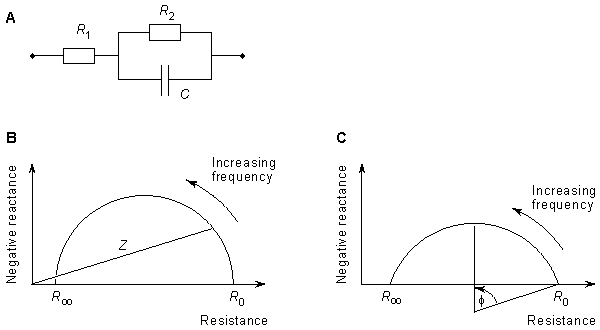
A useful method for illustrating the behavior of tissue impedance as a function of frequency is the Cole-Cole plot (Cole and Cole, 1941). In this presentation, real component R is plotted versus imaginary component X in the complex series impedance (R + jX) with the frequency as a parameter. Figure 25.1B shows the Cole-Cole plot of a three-element impedance with a single time constant, as shown in Figure 25.1A. The Cole-Cole plot obeys the following equation:

(25.3)
where Zf = impedance (as a function of frequency f ) R0 = resistance at f = 0 R 
= resistance at f = 
t = time constant (R2C) The Cole-Cole plot is a semicircle with radius (R 0 - R )/2 which intercepts the real axis at R 0 and R, a conclusion that can be verified by noting that the real (Re) and imaginary (Im) parts of Equation 25.3 satisfy

(25.4) )/2 with a radius of (R 0 - R )/2, as stated. In the three-element circuit of Figure 25.1A, R0 = R1 + R2, R = R 1 , and t = R2C.
In practice, the center of the semicircle is not necessarily on the real axis, but is located beneath it. The equation representing practical measurements may be described by Equation 25.4 (Schwan, 1957):

(25.5) The reactive component of human blood has been studied, for example, by Tanaka et al. (1970) and Zhao (1992). The reactive component of tissue impedance seems to have an important role in impedance plethysmography, as will be discussed later in this Chapter in connection with determining body composition..
(B) Cole-Cole plot for impedance with a single time constant.
(C) The depressed Cole-Cole plot.
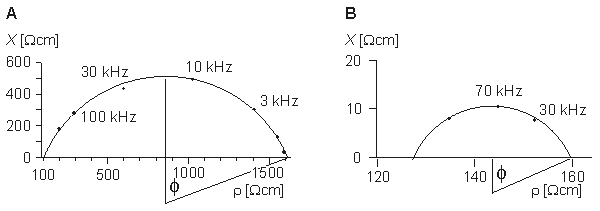
(B) longitudinal impedances of skeletal muscle.(Redrawn from Epstein and Foster, 1983.)
 | (25.6) |
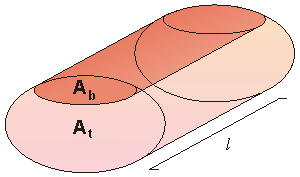
| where | Z | = longitudinal impedance of the model |
| Zb | = impedance of the blood volume | |
| Zt | = impedance of the tissue volume |
The relationship between the impedance change of the thorax and the impedance change of the blood volume is found by differentiating Equation 25.6 with respect to Zb:
 | (25.7) |
The impedance of the blood volume with blood resistivity rb based on the cylindrical geometry of Figure 25.5, is:
25.3.4 Determining the Stroke Volume
When determining stroke volume from thoracic impedance changes, Kubicek and colleagues (1966) and Kubicek (1968) made some assumptions concerning the relationship between stroke volume and net change in the thorax blood volume as evaluated in Equation 25.10. These assumptions are highly simplified and may be unreliable.
Assuming that Dt equals the ejection time te, DZ can be determined from equation

(25.8)
where rb = blood resistivity Ab = cross-section of the blood area l = length of the thorax model The relationship between changes in blood volume vb and the blood volume impedance is found by solving for the blood volume in Equation 25.8 and differentiating:

(25.9)
where vb = blood volume We finally derive the dependence of the change in blood volume on the change in thoracic impedance by solving for dZb in Equation 25.7 and substituting it into Equation 25.9:

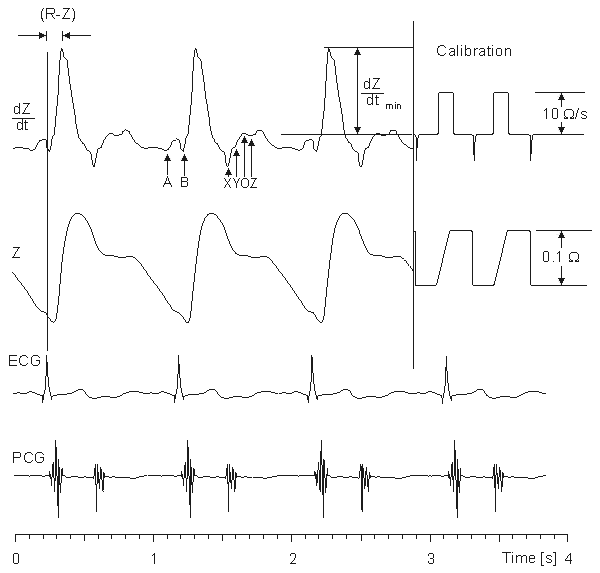
(25.10) As was mentioned earlier, during systole, the right ventricle ejects a volume of blood into the lungs. Subsequently, blood flows away from the lungs to the left atrium. The stroke volume can thus be determined from the impedance curve by extrapolating to the impedance (DZ), that would result if no blood were to flow out of the lungs during systole. (The underlying assumption is that DZ is determined mainly by changes in lung conductivity.)
In this extrapolation, it is assumed that if no blood were to flow away from the thorax during systole, the thorax impedance would continuously decrease during systole at a rate equal to the maximum rate of decrease of Z. Thus, DZ can be approximated graphically by drawing a tangent to the impedance curve at the point of its maximum rate of decrease, as illustrated in Figure 25.6. Then, the difference between the impedance values of the tangent line at the beginning and at the end of the ejection time is DZ.
The value of DZ is easy to determine with the help of the first derivative curve of the thoracic impedance signal. According to the definition of the derivative:

(25.11)

(25.12) With the above assumptions, the impedance change DZ can be determined by multiplying the ejection time by the minimum value of the first derivative of the impedance curve (that is, the maximum slope magnitude; the reader must remember that the slope is negative).
Finally, the formula for determining the stroke volume is obtained by substituting Equation 25.12 into Equation 25.10, which gives:

(25.13)
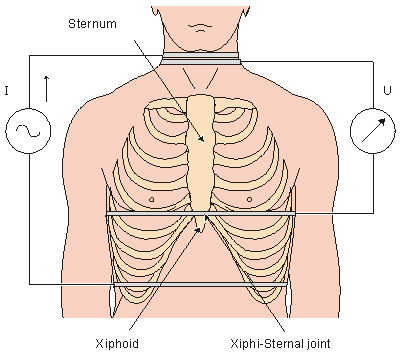
where SV = stroke volume [ml] rb = resistivity of the blood [W·cm] l = mean distance between the inner electrodes [cm] Z = mean impedance of the thorax [W] 
= absolute value of the maximum deviation of the first derivative signal during systole [W/s] te = ejection time [s] The ejection time can be determined from the first-derivative impedance curve with the help of the phonocardiogram or carotid pulse. Then, the impedance curve itself is used only for control purposes (e.g., checking the breathing).
The resistivity of the blood is of the order of 160 Wcm. Its value depends on hematocrit, as discussed in Section 7.4.
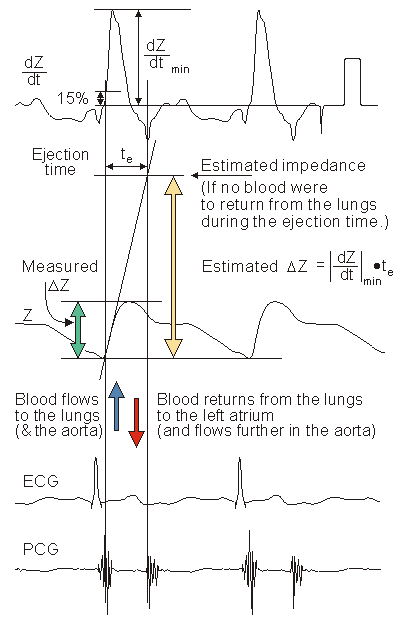
25.3.5 Discussion of the Stroke Volume Calculation Method
The method described above, developed by Kinnen and Kubicek, is widely used to estimate stroke volume from impedance recordings. We discuss later efforts to identify the source or sources of the measured changes in impedance. It will be seen that such research implicates changes in blood volume in the vena cava, atria, ventricles, aorta, thoracic musculature, and lungs. Obviously, the two-compartment model, above, is a gross simplification. Furthermore, the assumed cylindrical geometry is also a highly simplified approximation. And, finally, the change of blood conductivity with change in velocity has been entirely neglected in this model.
25.4 ORIGIN OF IMPEDANCE SIGNAL IN IMPEDANCE CARDIOGRAPHY
25.4.1 Model Studies
Kinnen et al. (1964c) constructed a cylindrical thorax model to investigate the origin of the impedance signal (see Figure 25.7). The inner cylinder represents the blood volume of the heart and the primary arteriovenous system of the thorax. The medium outside the inner cylinder represented the lungs. In this model, the computed resistance for the inner cylinder was 495 W and for the interspace 32 W. These values indicated that most of the current flux would tend to travel through the model's lungs so that the origin of the impedance signal should be based primarily on the right ventricle. This is consistent with observations in patients with septal defects (Lababidi et al., 1971). In these patients the cardiac output, measured by impedance plethysmography, correlates well with the blood flow in the pulmonary circulation.
25.4.2 Animal and Human Studies
Compared to the model studies, some practical experiments performed on animals gave different results concerning the origin of the signal. Baker, Hill, and Pale (1974) cite an experiment performed on a calf in which the natural heart was replaced by an implanted prosthesis containing artificial right and left ventricles. In this experiment the ventricles were operated either simultaneously or separately. The contribution of the left ventricle to the impedance signal was 62% of the total signal whereas that from the right ventricle was 38%.
25.4.3 Determining the Systolic Time Intervals from the Impedance Signal
Lababidi et al. (1970) carefully studied the timing of each significant notch in the first derivative curve of the thoracic impedance signal and assigned them to certain events in the heart cycle. According to their study, the relationship is as shown in Table 25.2 (see also Figure 25.3).
25.4.4 The Effect of the Electrodes
In impedance plethysmography, the current is fed from a constant current generator to the thorax by an electrode pair, and the voltage generated by this current is measured by another electrode pair. With a well-designed constant current generator the current in the thorax can be maintained constant despite electrode skin resistance changes. The mean impedance of the thorax is about 20 W. Consequently, the source impedance for the detected voltage is very low. If the voltmeter circuit is designed to have a high input impedance, the contact resistance can be neglected. In commercially manufactured equipment, the impedance is about 100 kW, in comparison to which the effects of contact impedance changes lie within an acceptable level (Kubicek, 1968).
25.4.5 Accuracy of the impedance cardiography
Today, more than one hundred publications exist on the accuracy of impedance cardiography. Lamberts, Visser, and Ziljstra (1984) have made an extensive review of 76 studies. In this chapter we discuss some representative studies where the accuracy of impedance cardiography has been evaluated. These can be divided into two main categories. In the first category the effect of the hematocrit on the blood resistivity is ignored and a constant value is used in the calculations for the resistivity of blood, usually 150 Wcm. In the second category, the value of the blood resistivity is first determined for each subject.
Experiments Where the Blood Resistivity is Constant
Kinnen and co-workers (1964b) determined the stroke volume from the equation
Experiments with Individual Resistivity Values
Lababidi et al. (1971) studied 95 children with various types of congenital heart disease using dye dilution and Fick principles as reference methods. In 20 subjects, paired impedance-dye dilution values had an average absolute difference of 6.6% ranging from -12% to +13% with a standard deviation of 0.259
Other Studies
Additional studies of the correlation between impedance methods and cardiac output reference techniques are summarized in Penney (1986). The results are generally similar to those described above. From these studies one can conclude that impedance cardiography is satisfactory for the determination of relative cardiac output for most normals. Under conditions of hypoxia, drugs, ventilatory maneuvers, and so on, the correlation may become poor.
25.5 OTHER APPLICATIONS OF IMPEDANCE PLETHYSMOGRAPHY
25.5.1 Peripheral Blood Flow
Impedance plethysmography is also a convenient method for conducting measurements of blood volume changes in applications other than cardiac stroke volume. The peripheral circulation can be studied by using an inflated cuff for blocking the venous flow and monitoring the blood volume increase in the limb. In such studies, Equation 25.9 is readily applicable (van de Water et al. 1971). Yamamoto, Yamamoto, and Öberg (1991 and 1992) have made technical and theoretical studies of the impedance plethysmography technique for measuring the blood flow in human limbs.
25.5.2 Cerebral Blood Flow
There are also applications where impedance plethysmography was used in an attempt to monitor cerebral blood flow. In these experiments one should be extremely careful in electrode placement to ensure that the impedance signal comes mainly from the intracranial region. As can be seen from the discussion in Section 13.4, even in the case where the bipolar leads in the inhomogeneous concentric spherical head model are located at opposite sides of the model, more than one third of the lead field current flows outside the skull. By moving the electrodes closer to each other, the relative amount of current outside the skull increases rapidly.
25.5.3 Intrathoracic Fluid Volume
Impedance plethysmography technique can also be used for monitoring intrathoracic fluids other than blood. The fluid in the pleural cavity has a considerable effect on the mean impedance of the thorax. Equation 25.9 can again be used to monitor the pleural fluid changes in the thorax (van de Water et al. 1971).
25.5.4 Determination of Body Composition
Bioelectric impedance may be used in determining the body composition. In this procedure the impedance is measured between one arm and one leg by feeding a current less than 1 mA rms at 50 kHz frequency. The determination of the body composition is based on measurement of the resistive and reactive components of the body impedance (Baumgartner, Chunlea, and Roche, 1988). With this method it is possible to estimate several parameters of the body composition such as total body water, fat free mass, body cell mass, and caloric consumption (Kushner and Shoeller, 1986; Lukaski et al., 1985). de Vries et al. (1989) have applied this technique for determining intracellular and extracellular fluid volumes during hemodialysis.
25.5.5 Other Applications
There have been some attempts to use the impedance technique to monitor cardiac contractility. Siegel et al. (1970), in an experiment with dogs, quantified myocardial contractility and the first-derivative thoracic impedance signal. Myocardial contractility and vascular tone was altered by the use of norepinephrine, isoproterenol, and methoxamine. They measured the time from the peak of the ECG R-wave to the peak first derivative of the isovolumic portion of the ventricular contraction (dp/dtmin) and to the inflection point in the first derivative of impedance (dZ/dt). From these curves they obtained a correlation of r = .88. This application has, however, not yet reached wide acceptance.
Sakamoto et al. (1979) constructed an anatomically more realistic model in which changes in vena cava, heart, lungs, aorta, and torso shape were investigated (see Figure 25.8). The model permits an examination of the effect of conductivity changes of component structures on the measured impedance. The weakness in this work is that one does not know what quantitative changes in conductivity are brought about as a result of real or simulated blood circulation. Sakamoto et al. (1979) also did studies with dogs and humans where they measured the isopotential lines on the surface of the thorax.
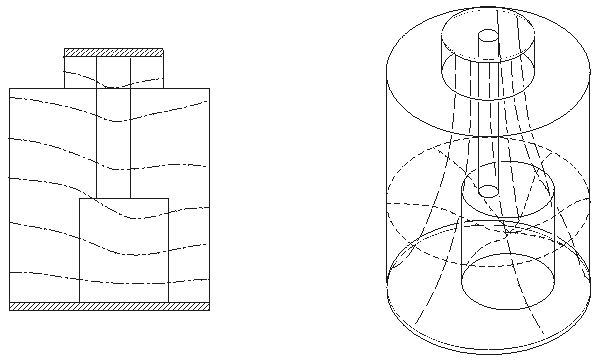
Fig. 25.7 Thorax model by Kinnen.
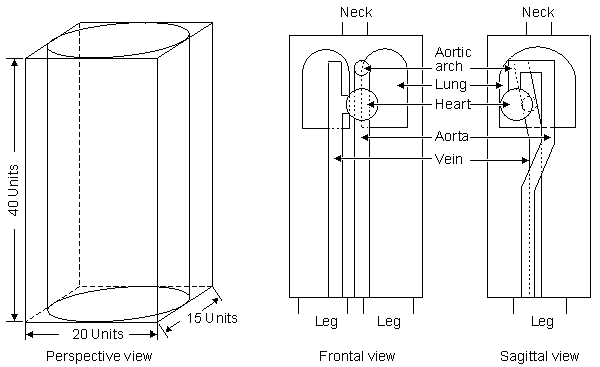
Fig. 25.8 Thorax model by Sakamoto et al.(1979).
Witsoe and Kottke (1967) conducted experiments with dogs, using venous occlusion achieved by an inflated ball. In these experiments the origin of the impedance signal was found to be contributed totally by the left ventricle. (This is also seen in humans.) Stroke volume measurements with impedance plethysmography on patients with aortic valve insufficiency give values that are too high.
Penney (1986) summarized a number of studies and estimated, on the base of these observations, the contributions to the impedance signal shown in Table 25.1.
Contributing organ Contribution Vena cava and right atrium +20% Right ventricle -30% Pulmonary artery and lungs +60% Pulmonary vein and left atrium +20% Left ventricle -30% Aorta and thoracic musculature +60% Source: Penney (1986) Mohapatra (1981) conducted a critical analysis of a number of hypotheses concerning the origin of the cardiac impedance signal. He concluded that it was due to cardiac hemodynamics only. Furthermore, the signal reflects both a change in the blood velocity as well as change in blood volume. The changing speed of ejection has its primary effect on the systolic behavior of DZ whereas the changing volume (mainly of the atria and great veins) affects the diastolic portion of the impedance curve.
These facts point out that the weakest feature of impedance plethysmography is that the source of the signal is not accurately known. Additional critical comments may be found in Mohapatra (1988).
impedance signal in impedance cardiography.
Event in the cardiac cycle Notch Atrial contraction A Closure of tricuspid valve B Closure of aortic valve X Closure of pulmonic valve Y Opening snap of mitral valve O Third heart sound Z Source: Lababidi et al., (1970) The first-derivative impedance curve can be used with some accuracy in timing various events in the cardiac cycle. The ejection time can be determined as the time between where the dZ/dt curve crosses the zero line after the B point, and the X point. However, in general, the determination of ejection time from the dZ/dt curve is more complicated. Thus, the need of the phonocardiogram in determining the ejection time depends on the quality and clarity of the dZ/dt curve. Though the timing of the various notches of the dZ/dt curve is well known, the origins of the main deflections are not well understood.
Hill, Jaensen, and Fling (1967) have introduced a critical comment concerning the effect of the contact impedance on the signal: they claim that the entire signal is an electrode artifact. Based on the preceding arguments and the experiments concerning the origin of the signal (Lababidi et al., 1971; Baker, Hill, and Pale, 1974) these claims can be ignored.
The effect of changes in the mean thoracic impedance has also been investigated (Hill and Lowe, 1973). Placement of a defibrillator back electrode under the back of a supine patient changed the mean impedance recorded by the instrument by up to 20%, but did not have any significant influence on the stroke volume value determined by the instrument, because of a simultaneous change in (dZ/dt)min, which compensated for the change in Z. This is easily seen by noting that stroke volume is proportional to Z-2, whereas dZ is proportional to Z2. Slight displacement of the detector electrodes changes the measured mean impedance and first derivative signal, but their effect on the computed stroke volume is compensated by the changed value of the mean distance of the electrodes. This is also easy to prove using the previous theory. It is also interesting to note that the signals remain unchanged when one half of the lower detector electrode is removed (Hill and Lowe, 1973). This implies that the electrode is situated on an equipotential surface, thus supporting the assumption of cylindrical symmetry.

(25.14)
where DZ = change of the impedance of the thorax Z = mean value of the impedance of the thorax vtx = volume of the thorax between the inner electrode pair They used the Fick principle as a reference for evaluating stroke volume. (The Fick principle determines the cardiac output from the oxygen consumption and the oxygen contents of the atrial and venous bloods.) In a study of six subjects at various exercise levels, the correlation between the impedance and Fick cardiac outputs was r = 0.962, with an estimated standard error of 12% of the average value of the cardiac output.
Harley and Greenfield (1968) performed two series of experiments with simultaneous dye dilution and impedance techniques. They estimated DZ from the impedance curve itself, instead of using the first-derivative technique. In the first experiment, 13 healthy male subjects were examined before and after an intravenous infusion of isoproterenol. The mean indicator dilution cardiac output was 6.3 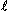 /min before and 9.5 /min after infusion. The ratios of the cardiac outputs measured with impedance plethysmography and indicator dilution were 1.34 and 1.23, respectively. This difference (p > .2) was not significant. The second experiment included 24 patients with heart disease, including aortic and mitral insufficiencies. A correlation coefficient of r = .26 was obtained for this data. The poor correlation was caused in those cases with aortic and mitral insufficiency.
Bache, Harley, and Greenfield (1969) performed an experiment with eight patients with various types of heart disease excluding valvular insufficiencies. As a reference they used the pressure gradient technique. Individual correlation coefficients ranged from .58 to .96 with an overall correlation coefficient as low as .28.
Baker et al. (1971) compared the impedance and radioisotope dilution values of cardiac output for 17 normal male subjects before and after exercise. The regression function for this data was COZ = 0.80·COI + 4.3 with a correlation coefficient r = .58. The comparison between the paired values before and after exercise showed better correlation for the impedance technique. Baker examined another group of 10 normal male subjects by both impedance and dye techniques. In 21 measurements the regression function was COZ = 1.06·COD + 0.52, with correlation coefficient r = .68. In addition to this set of data, the impedance cardiac output was determined by using individual resistivity values determined from the hematocrit. The relation between resistivity and Hct was, however, not mentioned.
In this case, the regression function was COZ = 0.96·COD + 0.56 with correlation coefficient r = .66. A set of measurements was performed also on 11 dogs using electromagnetic flowmeters and the impedance technique. A comparison of 214 paired data points was made with intravenous injections of epinephrine, norepinephrine, acetylcholine, and isoproterenol. Values of the correlation coefficients from each animal ranged from 0.58 to 0.98 with a mean value of 0.92. The first two experiments of this paper are also presented in Judy et al. (1969).
/min/m˛. Paired impedance-cardiac output values had an absolute difference of 3.1%, ranging from -15% to +3.2% with a standard deviation of 0.192 /min/m˛. The F-test showed the reproducibility of both methods to be similar: F = 1.82 and p > .05. For 53 sequential determinations of impedance cardiac output and dye dilution, the absolute mean difference was -1.8%, t = 1.19 and p &"62; .05. When determining, sequentially, the relationship between Fick and dye dilution principles, 37 of 39 points fell within 20% limits. The absolute mean difference was 8.3%, and the algebraic mean difference was +3.4%. The correlation between impedance and Fick cardiac outputs was r = .97. These studies were performed with patients without intracardiac shunts or valvular insufficiencies.
A comparison of impedance cardiac output to Fick systemic cardiac output in patients with left to right shunts showed the correlation to be poor: r = .21. However, a comparison of the impedance cardiac output to the Fick pulmonary blood flow in these cases gave a correlation of r = .96 (see Fig. 25.9).
Baker, Hill, and Pale (1974) compared impedance and dye dilution cardiac outputs in three dogs and got a correlation of r = .879.
Malmivuo (1974) compared impedance and Fick methods in 18 patients without valvular incompetencies, but with one subject having a left to right shunt. For this special subject a comparison was made to pulmonary blood flow. The regression function was COZ = 0.97·COF + 0.45 yielding a correlation coefficient of r = .97 (see Figure 25.8).
Malmivuo, Orko, and Luomanmäki (1975) compared impedance and Fick methods in 11 patients with atrial fibrillation and without intracardiac shunts or valvular insufficiencies. The regression function was COZ = 1.05·COF + 0.1, with a correlation coefficient of r = .96.
/min before and 9.5 /min after infusion. The ratios of the cardiac outputs measured with impedance plethysmography and indicator dilution were 1.34 and 1.23, respectively. This difference (p > .2) was not significant. The second experiment included 24 patients with heart disease, including aortic and mitral insufficiencies. A correlation coefficient of r = .26 was obtained for this data. The poor correlation was caused in those cases with aortic and mitral insufficiency.
Bache, Harley, and Greenfield (1969) performed an experiment with eight patients with various types of heart disease excluding valvular insufficiencies. As a reference they used the pressure gradient technique. Individual correlation coefficients ranged from .58 to .96 with an overall correlation coefficient as low as .28.
Baker et al. (1971) compared the impedance and radioisotope dilution values of cardiac output for 17 normal male subjects before and after exercise. The regression function for this data was COZ = 0.80·COI + 4.3 with a correlation coefficient r = .58. The comparison between the paired values before and after exercise showed better correlation for the impedance technique. Baker examined another group of 10 normal male subjects by both impedance and dye techniques. In 21 measurements the regression function was COZ = 1.06·COD + 0.52, with correlation coefficient r = .68. In addition to this set of data, the impedance cardiac output was determined by using individual resistivity values determined from the hematocrit. The relation between resistivity and Hct was, however, not mentioned.
In this case, the regression function was COZ = 0.96·COD + 0.56 with correlation coefficient r = .66. A set of measurements was performed also on 11 dogs using electromagnetic flowmeters and the impedance technique. A comparison of 214 paired data points was made with intravenous injections of epinephrine, norepinephrine, acetylcholine, and isoproterenol. Values of the correlation coefficients from each animal ranged from 0.58 to 0.98 with a mean value of 0.92. The first two experiments of this paper are also presented in Judy et al. (1969).
/min/m˛. Paired impedance-cardiac output values had an absolute difference of 3.1%, ranging from -15% to +3.2% with a standard deviation of 0.192 /min/m˛. The F-test showed the reproducibility of both methods to be similar: F = 1.82 and p > .05. For 53 sequential determinations of impedance cardiac output and dye dilution, the absolute mean difference was -1.8%, t = 1.19 and p &"62; .05. When determining, sequentially, the relationship between Fick and dye dilution principles, 37 of 39 points fell within 20% limits. The absolute mean difference was 8.3%, and the algebraic mean difference was +3.4%. The correlation between impedance and Fick cardiac outputs was r = .97. These studies were performed with patients without intracardiac shunts or valvular insufficiencies.
A comparison of impedance cardiac output to Fick systemic cardiac output in patients with left to right shunts showed the correlation to be poor: r = .21. However, a comparison of the impedance cardiac output to the Fick pulmonary blood flow in these cases gave a correlation of r = .96 (see Fig. 25.9).
Baker, Hill, and Pale (1974) compared impedance and dye dilution cardiac outputs in three dogs and got a correlation of r = .879.
Malmivuo (1974) compared impedance and Fick methods in 18 patients without valvular incompetencies, but with one subject having a left to right shunt. For this special subject a comparison was made to pulmonary blood flow. The regression function was COZ = 0.97·COF + 0.45 yielding a correlation coefficient of r = .97 (see Figure 25.8).
Malmivuo, Orko, and Luomanmäki (1975) compared impedance and Fick methods in 11 patients with atrial fibrillation and without intracardiac shunts or valvular insufficiencies. The regression function was COZ = 1.05·COF + 0.1, with a correlation coefficient of r = .96.
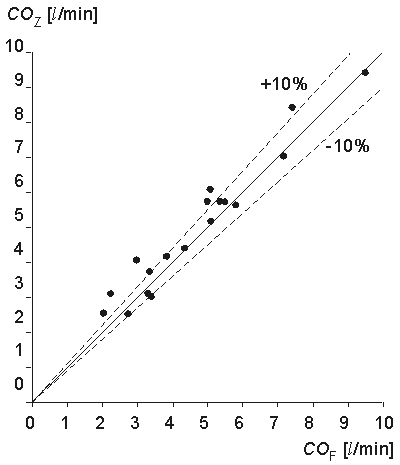
Fig. 25.9 Comparison of the impedance and Fick methods in determining the pulmonary blood flow.
In evaluating the significance of a particular correlation coefficient between impedance and reference methods, Penney points out that the reference methods themselves are not completely consistent. For example, if one considers the correlation coefficient r, then between Fick and dye dilution .95 < r < .999; Fick and thermodilution .70 < r < .99; Fick to carbon dioxide breathing, r = .94; dye to thermodilution, .68 < r < .99.
Concerning the accuracy of impedance plethysmography in determining peripheral blood flow, there are much fewer data available as few detailed experiments have been published. van de Water et al.(1971) reported on a series of measurements in a hind limb of a dog using an electromagnetic flowmeter as a reference method. A correlation of r = .962 was obtained, using a constant value for the resistivity of the blood.
From this it is easy to deduce that if the impedance measurement is made with electrodes placed on one side of the head only or if, when using circular band electrodes, the electrodes are relatively close to each other, the major part of the signal comes from the blood flow in the scalp, not from that in the brain area. This shading effect of the skull does not show up as clearly in the EEG-measurement, because no bioelectric sources exist outside the skull (Malmivuo, 1992).
Van de Water et al. (1971) infused 400 cmł of saline in 25 cmł increments into a thoracic cavity of a dog weighing 15 kg. The regression formula between the infused saline volume and the thoracic impedance was Z = 0.02281 cmł + 46.944 with a correlation coefficient of r = .988. They reported also one case when 900 cmł of pleural fluid was removed from a patient in 50 cmł increments. The regression formula in this case was Z = 0.0024 cmł + 17.57, with a correlation coefficient of r = .965.