B.5 MAGNETIC VECTOR POTENTIAL AND ELECTRIC SCALAR POTENTIAL IN THE REGION OUTSIDE THE SOURCES
PRECONDITIONS:
SOURCE: Quasistatic  i (w < 1000 Hz)
i (w < 1000 Hz)
CONDUCTOR: Limited finite (r < 1 m) region outside the sources inhomogeneous resistive (we/s < 0.15), µ = µ0, e = e0
In this section we derive from Maxwell's equations the equations for magnetic vector potential 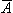 and electric scalar potential F in physiological applications, Equations B.19 and B.21, respectively.
and electric scalar potential F in physiological applications, Equations B.19 and B.21, respectively.
 Since the divergence of
Since the divergence of 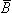 is identically zero (Equation B.9), the magnetic field may be derived from the curl of an arbitrary vector field , which is called the magnetic vector potential. This fulfills the requirement stated in Equation B.9 because the divergence of the curl of any vector field is necessarily zero. Consequently,
is identically zero (Equation B.9), the magnetic field may be derived from the curl of an arbitrary vector field , which is called the magnetic vector potential. This fulfills the requirement stated in Equation B.9 because the divergence of the curl of any vector field is necessarily zero. Consequently,
  | (B.12) |
Since = m0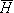 , we can substitute Equation B.12 into Equation B.6. We consider only the volume conductor region external to the membranes where the emfs are zero (note that the emfs are explicitly included within the membrane in the form of Nernst potential batteries), and we consequently obtain
, we can substitute Equation B.12 into Equation B.6. We consider only the volume conductor region external to the membranes where the emfs are zero (note that the emfs are explicitly included within the membrane in the form of Nernst potential batteries), and we consequently obtain
 | (B.13) |
Now, when the curl of a vector field is zero, that vector field can be derived as the (negative) gradient of an arbitrary scalar potential field (which we designate with the symbol F and which denotes the electric scalar potential). This assignment is valid because the curl of the gradient of any scalar field equals zero. Thus Equation B.13 further simplifies to
 | (B.14) |
According to the Helmholtz theorem, a vector field is uniquely specified by both its divergence and curl (Plonsey and Collin, 1961). Since only the curl of the vector field has been specified so far (in Equation B.12), we may now choose
 | (B.15) |
This particular choice eliminates F from the differential equation for (Equation B.17). That is, it has the desirable effect of uncoupling the magnetic vector potential from the electric scalar potential F. Such a consideration was originally suggested by Lorentz when dealing with the free-space form of Maxwell's equations. Lorentz introduced an equation similar to Equation B.15 known as the Lorentz condition, which is that
 | (B.16) |
We have modified this expression since we have eliminated in Equations B.10 and B.11 the displacement current jwe in favor of a conduction current s. This amounts to replacing jwe by s in the classical Lorentz condition (Equation B.16), resulting in Equation B.15. The Lorentz condition can also be shown to have another important property, namely that it ensures the satisfaction of the continuity condition.
in favor of a conduction current s. This amounts to replacing jwe by s in the classical Lorentz condition (Equation B.16), resulting in Equation B.15. The Lorentz condition can also be shown to have another important property, namely that it ensures the satisfaction of the continuity condition.
Now, if we substitute Equations B.12, B.14, and B.15 into Equation B.10, keeping in mind that = /m0, and if we use the vector identity that
 | (B.17) |
we obtain
 | (B.18) |
Just as emfs were eliminated by confining attention to the region external to the excitable cell membranes, so too could one eliminate the nonconservative current i in Equation B.10. In this case all equations describe conditions in the passive extracellular and intracellular spaces; the effect of sources within the membranes then enters solely through boundary conditions at and across the membranes. On the other hand, it is useful to retain i as a distributed source function in Equation B.10. While it is actually confined to cell membranes ensuring the aforementioned boundary conditions, it may be simplified (averaged) and regarded as an equivalent source that is uniformly distributed throughout the "source volume." For field points outside the source region which are at a distance that is large compared to cellular dimensions (over which averaging of i occurs) the generated field approaches the correct value.
Equation B.18 is known as the vector Helmholtz equation, for which solutions in integral form are well known in classical electricity and magnetism (Plonsey and Collin, 1961). Adapting such a solution to our specific equation gives
 | (B.19) |
where
Note that r is the radial distance from a source element dV(x,y,z) (unprimed coordinates) to the field point P(x',y',z') (primed coordinates), and is thus a function of both the unprimed and primed coordinates.
To evaluate an upper bound to the magnitude of kr in the exponential terms in Equation B.19 we choose:
rmax = 100 cm
w = 2p·1000 1/s
µ0 = 4p·10-9 H/cm
s = .004 S/cm
Then
Since e-.04 = .96, these exponential terms can be ignored and we get a simplification for Equation B.19, giving the magnetic vector potential under electrophysiological conditions:
 | (B.20) |
The electric scalar potential F may be found from by using Equation B.15 with Equation B.20. In doing so, we note that Equation B.20 involves an integration over the source coordinates (x,y,z) while Equation B.15 involves operations at the field coordinates (x',y',z'). Consequently, we get
 | (B.21) |
where 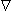 ' operates only on the field coordinates, which is why i is not affected. Since '(1/r) = –(1/r), we finally get for the electric scalar potential:
' operates only on the field coordinates, which is why i is not affected. Since '(1/r) = –(1/r), we finally get for the electric scalar potential:
 | (B.22) |
Equation B.22 is identical to static field expressions for the electric field, where i is interpreted as a volume dipole density source function. This equation corresponds exactly to Equation 7-5. Although a staticlike equation applies, i is actually time-varying, and consequently, so must F be time-varying synchronously. We call this situation a quasistatic one.
When the source arises electrically (including that due to cellular excitation), a magnetic field is necessarily set up by the resulting current flow. The latter gives rise to a vector potential , which in turn contributes to the resulting electric field through the term jw in Equation B.14. However, under the conditions specified, |w| is negligible compared to the term |F| as discussed in Plonsey and Heppner (1967). Under these conditions we are left with the scalar potential term alone, and Equation B.14 simplifies to
 | (B.23) |
which also corresponds to a static formulation. It should be kept in mind that Equation B.23 is not exact, but only a good approximation. It corresponds to the quasistatic condition where the electric field resembles that arising under static conditions. Under truly static conditions the electric and magnetic fields are completely independent. Under quasistatic conditions, while the fields satisfy static equations, a low frequency time variation may be superimposed (justified by the low frequency conditions discussed earlier), in which case the magnetic field effects, although extant, can normally be ignored.
Note that in this case, where the sources are exclusively bioelectric and the simplification of Equation B.23 is valid, Equation B.11 leads to Equation 7.2 ( = i – sF).
B.6 STIMULATION WITH ELECTRIC AND MAGNETIC FIELDS
B.6.1 Stimulation with Electric Field
PRECONDITIONS:
SOURCE: Steady-state electric field
CONDUCTOR: Uniform fiber in volume conductor
The above comments notwithstanding, we are also interested in a situation where excitable tissue is stimulated solely with an applied magnetic field. In this case the vector potential is large and cannot be ignored. In fact, to ignore under these circumstances is to drop the underlying forcing function, which would leave an absurd result of no field, either electric or magnetic.
We have shown in Chapter 3 that for a single uniform fiber under steady-state conditions a homogeneous partial differential equation (Equation 3.46) arises:
 | (B.24) |
| where | Vm | = transmembrane potential |
| | l | = space constant, characteristic of the physical and electric properties of the fiber |
| | x | = coordinate along the direction of the fiber |
For a point source at the origin we have also essentially shown, in Chapter 3, that the solution to Equation B.24 is (Equation 3.49)
 | (B.25) |
where V'(x) = deviation of the membrane voltage from the resting voltage.
In this equation
 | (B.26) |
| where | Vm(0) | = transmembrane potential at the origin |
| | I0 | = applied intracellular point current |
| | ri | = intracellular axial resistance per unit length |
We remark, here, that for a more general applied scalar potential field, Fe, Equation B.24 becomes
 | (B.27) |
One can recognize in this equation that the second derivative of the applied potential field along the fiber is the forcing function (in fact, it has been called the "activating function"), whereas the dependent variable, Vm, is the membrane response to the stimulation. Using Equation B.23, one can write Equation B.24 as
 | (B.28) |
where is the applied electric field.
B.6.2 Stimulation with Magnetic Field
PRECONDITIONS:
SOURCE: Time-varying magnetic field
CONDUCTOR: Uniform fiber in volume conductor
Electric stimulation may be produced by applying a time-varying magnetic field to the tissue. As given in Equation B.12, this magnetic field is defined as the curl of a vector potential . Now the stimulus is introduced solely through a magnetic field that induces an electric field . Equation B.27 is still completely valid except that the applied field is found from Equation B.14, namely where = –jw.
The determination of the vector field from a physical coil is found, basically, from Equation B.20 (which corresponds to Equation 12.33). This relationship has also been worked out and published for many other coil configurations.
We also note that since the differential equations B.24, B.27, and B.28 are linear, and the solution given in Equation B.25 is essentially the response to a (spatial) unit impulse at the origin (set I0 = d(x)), then linear systems theory describes the solution to Equation B.27, (or B.28), as
 | (B.29) |
where Ä denotes convolution. (The added factor of rm is required in order to convert the right side of Equation B.29 into a current density.) The convolution operation can be performed by taking the inverse Fourier transform of the product of the Fourier transform of V' and the Fourier transform of the second derivative of de. Such operations are readily carried out using the fast Fourier transform (FFT).
B.7 SIMPLIFIED MAXWELL'S EQUATIONS IN PHYSIOLOGICAL PREPARATIONS IN THE REGION OUTSIDE THE SOURCES
PRECONDITIONS:
SOURCES and FIELDS: Quasistatic (w < 1000 Hz) i, ,
CONDUCTOR: Limited finite (r < 1 m) region outside the sources inhomogeneous resistive (we/e < 0.15) µ = µ0, e = e0
We finally collect the Maxwell's equations in their simplest form. These equations are valid under quasistatic electrophysiological conditions outside the region of bioelectric sources:
| (B.23) |
 | (B.10) |
 | (B.11) |
 | (B.09) |
REFERENCES
Barr RC, Spach MS (1977): Sampling rates required for digital recording of intracellular and extracellular cardiac potentials. Circulation 55: 40-8.
Maxwell J (1865): A dynamical theory of the electromagnetic field. Phil. Trans. R. Soc. (Lond.) 155: 459-512.
Maxwell J (1873): Treatise On Electricity and Magnetism, Vol. 2, Oxford. (Reprint by Dover, New York, 1954.)
Plonsey R, Collin R (1961): Principles and Applications of Electromagnetic Fields, 554 pp. McGraw-Hill, New York.
Plonsey R, Heppner DB (1967): Considerations of quasistationarity in electrophysiological systems. Bull. Math. Biophys. 29:(4) 657-64.
Schwan HP, Kay CF (1957): The conductivity of living tissue. Ann. N.Y. Acad. Sci. 65: 1007.








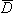 to the sources that generate it, namely the charge density r.
to the sources that generate it, namely the charge density r. 



 0.
0.
