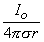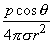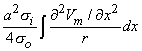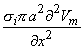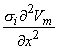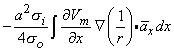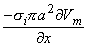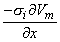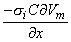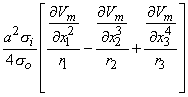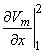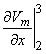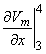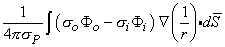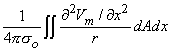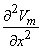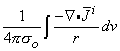8.2 SOURCE MODELS
8.2.1 Monopole
PRECONDITIONS:
Source: Monopole in a fixed location
Conductor: Infinite, homogeneous
The simplest source configuration is the point source or monopole. If we consider a point current source of magnitude I0 lying in a uniform conducting medium of infinite extent and conductivity s, then current flow lines must be uniform and directed radially. As a consequence, for a concentric spherical surface of arbitrary radius r, the current density J crossing this surface must be uniform and will equal I0 divided by the total surface area. That is
 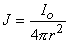 | (08.01) |
since the total current is conserved. Because the current is everywhere in the radial direction, the current density expressed as a vector is
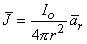 | (08.02) |
where  r = unit vector in the radial direction, where the origin is at the point source.
r = unit vector in the radial direction, where the origin is at the point source.
 Associated with the current flow field defined by Equation 8.2 is a scalar potential field F. Since the field is everywhere radial, there should be no variation of potential along a transverse direction, namely that on which r is a constant. Consequently, we expect isopotential surfaces to be a series of concentric spheres surrounding the point source with diminishing potentials for increasing values of r. In a formal sense, it is known from field theory that the electric field
Associated with the current flow field defined by Equation 8.2 is a scalar potential field F. Since the field is everywhere radial, there should be no variation of potential along a transverse direction, namely that on which r is a constant. Consequently, we expect isopotential surfaces to be a series of concentric spheres surrounding the point source with diminishing potentials for increasing values of r. In a formal sense, it is known from field theory that the electric field  is related to a scalar potential F by
is related to a scalar potential F by
From Ohm's law it follows that
Applying Equations 8.3 and 8.4 to 8.2 results in
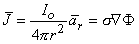 | (08.05) |
To satisfy Equation 8.5, only the component of 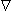 F in the direction of r can arise. This leads to
F in the direction of r can arise. This leads to
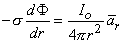 | (08.06) |
and integration with respect to r leaves us with
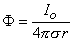 | (08.07) |
As suspected above, F is a constant on surfaces where r is constant (i.e., concentric spheres). Normally the potential for r 
 is set to zero, which accounts for having chosen the constant of integration in Equation 8.7 equal to zero. We note from Equation 8.7 that equipotential surfaces are indeed concentric spheres and that the potential magnitude is inversely proportional to the radius (with the origin at the monopole source).
is set to zero, which accounts for having chosen the constant of integration in Equation 8.7 equal to zero. We note from Equation 8.7 that equipotential surfaces are indeed concentric spheres and that the potential magnitude is inversely proportional to the radius (with the origin at the monopole source).
It is not always convenient to place the coordinate system origin at the point source (e.g., when considering several such sources). In this case it is desirable to distinguish the coordinates of the point source(s) from that of the field point, and we do this by using primes for the field point coordinates. Equation 8.7 then applies with, r given by
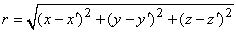 | (08.08) |
where each monopole is located at (x, y, z) while the field point is at (x', y', z').
The field described by Equation 8.7 for a point current source is identical to the electrostatic field from a point charge, provided that I0 is replaced by Q0 (the charge magnitude), s is replaced by e (the permittivity), and  replaced by . This result is not surprising since if the aforementioned exchanges are made, the governing equations for current flow convert exactly into those for electrostatics. This means that simply by interchanging symbols, solutions to problems in electrostatics can be converted into solutions to equivalent problems in current flow (and vice versa).
replaced by . This result is not surprising since if the aforementioned exchanges are made, the governing equations for current flow convert exactly into those for electrostatics. This means that simply by interchanging symbols, solutions to problems in electrostatics can be converted into solutions to equivalent problems in current flow (and vice versa).
The aforementioned is an example of duality. It can be a useful tool when there is an extensive literature already in existence. Sometimes there may be a limitation in physically realizing a condition in one or the other dual systems. For example, one can have zero conductivity, but the permittivity can never be less than that of vacuum. Also, while one can have a point charge, one cannot actually have a physical point source.
The reader may wonder why there is an interest in a point current source when such is not physically obtainable. One reason is that in a limited region, the fields may behave as if they arise from such a source (we say that the source is equivalent). Second, one can actually have two point sources of opposite polarity, in which case the field of interest can be found by the superposition of point source fields. In fact, this very situation is examined in the next section..8.2.2 Dipole
PRECONDITIONS:
Source: Dipole in a fixed location
Conductor: Infinite, homogeneous
In bioelectricity one can never have a single isolated monopole current source because of the need to conserve charge. But collections of positive and negative monopole sources are physically realizable if the total sum is zero. The simplest collection, and one that reflects a fundamental bioelectric source, is the dipole. The dipole consists of two monopoles of opposite sign but equal strength I0 (often termed source and sink) separated by a very small distance, d. In fact, the strict definition requires d 0, I0 with p = I0d remaining finite in the limit. The quantity p is the dipole moment or dipole magnitude. The dipole is a vector whose direction is defined from the negative point source to the positive. In fact, if  is the displacement from negative to positive point source and d a unit vector in that direction, then
is the displacement from negative to positive point source and d a unit vector in that direction, then
where 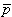 = the dipole vector.
= the dipole vector.
A dipole of arbitrary orientation is illustrated in Figure 8.1, where the coordinate system origin is placed at the negative pole. If the positive pole were also at the origin, the sources would cancel each other and their field would be zero. Consequently, the field arising from the displacement of the positive pole from the origin to its actual position (shown in Figure 8.1) is, in fact, the dipole field. But this can be found by examining the expression describing the potential of the positive monopole and evaluating the change in potential brought about by moving the monopole from the origin to its dipole position. And this, in turn, can be approximated from the first derivative of the monopole's potential field with respect to the source coordinates evaluated at the origin (as in a Taylor series representation). Specifically, to obtain the dipole field, a derivative of F (as given in Equation 8.7) is taken with respect to the direction (a directional derivative) and then multiplied by the magnitude of d. Thus, denoting the dipole field Fd, and based on Equation 8.7, we have
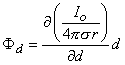 | (08.10) |
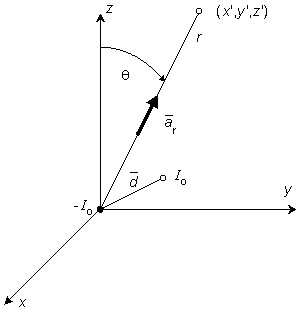
Figure 8.1. Dipole consisting of a sink -I0 at origin and a source I0 at radius vector , where d 0. Also illustrated is a field point at radius vector rr and polar (colatitude) angle q.
The directional derivative in Equation 8.10 equals the component of the gradient in the direction so that
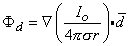 | (08.11) |
and, finally since I0d = p
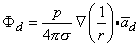 | (08.12) |
The accuracy of Equation 8.10 improves as d 0, and in fact, p (as noted earlier) is normally defined in the limit that d 0, I , such that the product I0d is finite and is equal to p. Consequently, Equation 8.12 is a rigorous (exact) expression for a mathematically defined dipole.
If the coordinate axes are oriented so that the dipole is directed along z- (the polar) axis and the dipole is placed at the origin, then carrying out the gradient operation in Equation 8.12 and noting that
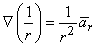 | (08.13) |
where r is oriented from the source point to field point, we obtain for the field of a dipole
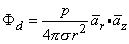 | (08.14) |
and
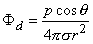 | (08.15) |
In Equation 8.15 the angle q is the polar (colatitude) angle. The above expressions can be confirmed by noting that the gradient operator (in Equation 8.13) acts on the source (unprimed) coordinates in Equation 8.8.
A comparison of the dipole field to a monopole field, by contrasting Equation 8.15 with Equation 8.7, shows that the dipole field varies as (1/r)2 whereas the monopole field varies as (1/r). In addition, the dipole equipotential surfaces are not concentric spheres but, rather, are more complicated, because of the factor cosq. The maximum dipole potential, for a given value of r, is on the polar axis (z axis).
8.2.3 Single Isolated Fiber: Transmembrane Current Source
PRECONDITIONS:
Source: Active fiber of finite or infinite length with circular cross-section
Conductor: Infinite, homogeneous
Figure 8.2 illustrates a long, thin excitable fiber lying in a uniform conducting medium of conductivity so and of unlimited extent. If we assume the existence of a propagating nerve impulse, then the activation currents are associated with a transmembrane current distribution im(x). Since the fiber is very thin and there is axial symmetry, we can describe the transmembrane current as a function of the axial variable x only. Thus the source description is one-dimensional. The dimension of im(x) is current per unit length. A small element of current im(x)dx can, therefore, be considered to behave like a point current source (a monopole) within the extracellular medium. Consequently, from Equation 8.7, we have
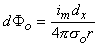 | (08.16) |
where r is given by Equation 8.8, Fo is the potential field and so is the conductivity outside the fiber (i.e.,extracellular conductivity). Integration over the fiber (i.e., with respect to x) gives the total field as
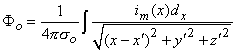 | (08.17) |
where the source is assumed to lie on the fiber axis, at (x, 0, 0), and the (fixed) field point is at (x', y', z').
We may apply the equations derived in Chapter 3, Section 3.4.2, to the fiber in Figure 8.2. We may approximate that the resistance of the interstitial medium ro 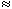 0 and that similarly the potential in the interstitial medium Fo 0. Using these approximations and Equation 3.42 and noting that Fi - Fo Vm we obtain
0 and that similarly the potential in the interstitial medium Fo 0. Using these approximations and Equation 3.42 and noting that Fi - Fo Vm we obtain
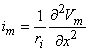 | (08.18) |
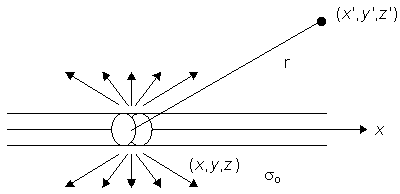
Figure 8.2. A long thin fiber is shown embedded in a uniform conducting medium of conductivity so and infinite in extent. The transmembrane current density is described by im(x) so that im(x)dx, illustrated, behaves as a point source in the extracellular medium.
so that Equation 8.17 may be written



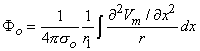
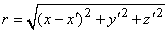
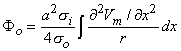
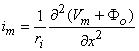
 Vm/
Vm/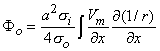
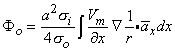
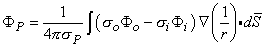
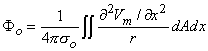
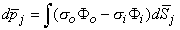
 1010 cells of which perhaps 5% are active at any moment during depolarization, the number of dipole source elements is extremely high. Under these conditions one can define a volume dipole moment density function (i.e., a dipole moment per unit volume) by averaging the dipole elements in each small volume. That is,
1010 cells of which perhaps 5% are active at any moment during depolarization, the number of dipole source elements is extremely high. Under these conditions one can define a volume dipole moment density function (i.e., a dipole moment per unit volume) by averaging the dipole elements in each small volume. That is,
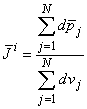
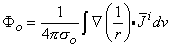
 (
(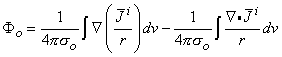
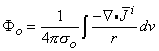
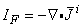
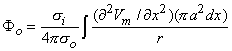
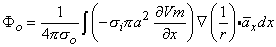
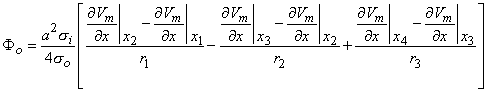
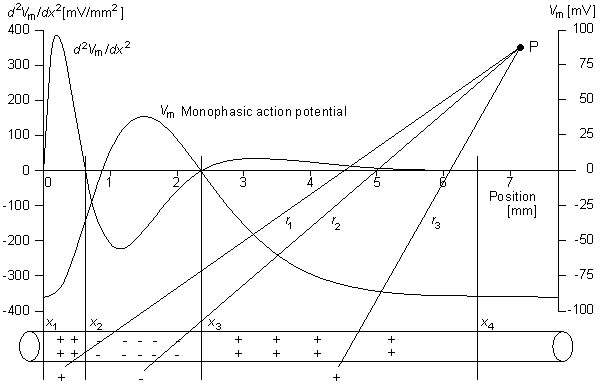
 0) are assumed to be planar. All fibers in the bundle are assumed to be parallel and carrying similar action potentials; consequently, each fiber will contain a similar equivalent source density. This is shown as a dipole density and hence proportional to -
0) are assumed to be planar. All fibers in the bundle are assumed to be parallel and carrying similar action potentials; consequently, each fiber will contain a similar equivalent source density. This is shown as a dipole density and hence proportional to -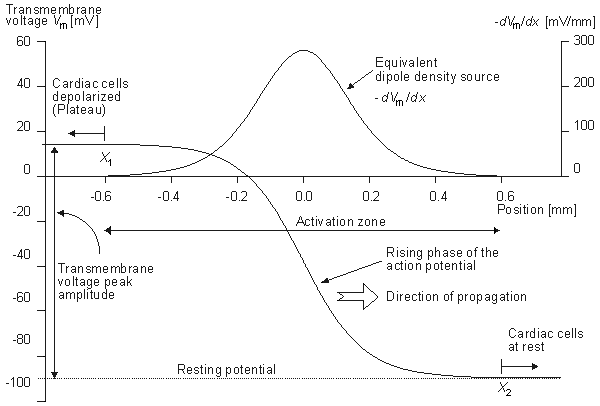
 in Equation 8.28 may be assumed to be constant in the integration over each cell. Thus each cell can be thought of as contributing a single lumped dipole source, which is simply the vector sum of its double layer surface elements. That is, the dipole for the jth cell, d
in Equation 8.28 may be assumed to be constant in the integration over each cell. Thus each cell can be thought of as contributing a single lumped dipole source, which is simply the vector sum of its double layer surface elements. That is, the dipole for the jth cell, d